Polymorfismy v genovém clusteru interleukinu 1: rizikový faktor alkoholické jaterní cirhózy
Jan Petrášek Orcid.org , Soňa Režnáková1, Jaroslav Hubáček Orcid.org , Jan Šperl Orcid.org 2, Pavel Trunečka3, Julius Špičák Orcid.org 2, Milan Jirsa Orcid.org 4
+ Pracoviště
Souhrn
Cíl studie: V patogenezi alkoholické jaterní cirhózy (ALC) hrají významnou úlohu zánětlivé cytokiny tumor necrosis factor alfa (TNF-ct) a interleukin 1-beta (IL-1 (3). Promotorové polymorfismy TNF-A -863C/A, TNF-A -238G/A, haplotyp [-511C; -31T] promotoru genu IL-1B a variabilní počet tandemových repetic délky 86-bp ve 2. intronu genu pro receptorového antagonistu IL-1 (IL-1RN) zvyšují transkripci genů TNF-A resp. IL-1B. Cílem studie bylo ověřit, zda tyto polymorfismy ovlivní riziko ALC.
Materiál a metodika: Frekvence alel uvedených polymorfismů byly stanoveny u 100 pacientů s etylickou cirhózou a u 180 neabstinujících zdravých dobrovolníků se srovnatelným zastoupením věku a pohlaví.
Výsledky: Žádná z alel TNF-A -863A, TNF-A -238A, IL-1B -511C, IL-1B -31T aILl-RN*2 nebyla samostatně asociována s ALC. Riziko ALC však bylo zvýšené u homozygotů pro alelu IL-1RN*2, kteří byli současně nosiči alely IL-1B -31T (relativní riziko 8,3, 95% konfidenční interval 1,7-40,7).
Závěr: Výsledky prokazují zvýšené riziko ALC u homozygotů pro alelu IL-1RN*2, kteří jsou alespoň heterozygoty pro alelu IL-1B -31T. Synergický účinek alel IL-1B -31T a IL-1RN*2 zvyšuje produkci IL-1β, který vyvolá polymorfonukleární infiltrát v játrech, působí profibrogenně a vysvětluje tak biologickou podstatu zvýšeného rizika ALC.
Klíčová slova: alkoholická jaterní cirhóza - genetický polymorfismus - interleukin-1 beta - neutrofilní granulocyty - tumor necrosis factor-alfa.
ÚVOD
Alkoholická jaterní cirhóza (ALC) představuje více než 50% chronických onemocnění jater a v rozvinutých zemích je příčinou kolem 50 000 úmrtí ročně (1,2) . Studie na dvojčatech prokázaly, že individuální citlivost jater vůči alkoholu je dědičně podmíněna (3,4) , avšak genetické faktory ovlivňující riziko rozvoje jaterní cirhózy u alkoholiků nebyly identifikovány (2) .
V patogenezi ALC se kromě přímého působení etanolu uplatňuje i zánětlivá odpověď jater na bakteriální endotoxiny (lipopolysacharid, LPS). Etanol zvyšuje střevní permeabilitu pro LPS, který je transportován do jater, kde aktivuje Kupfferovy buňky (KC) (5) produkující tumor necrosis factor a (TNF-a) a interleukin 1β (IL-1β). Tumor necrosis factor a indukuje cestou TNF-a receptoru I. typu apoptózu hepatocytů u akutní alkoholické steato-hepatitidy a jeho koncentrace v krvi koreluje s krátkodobou mortalitou (6,7) . Interleukin 1β, jehož receptor intracelulární apoptotickou doménu nemá, je hlavním cytokinen iniciujícím infiltraci jater polymorfonukleárními leukocyty (PMN), které převažují v zánětlivém infiltrátu alkoholické nemoci jater (8,9) . Kontinuální expozice jaterního parenchymu lyzosomálním enzymům z rozpadlých granulocytů vede ke chronickému poškozování hepatocytů, aktivaci hvězdicových buněk a k rozvoji fibrózy (10,11) . Oba cytokiny přispívají k aktivaci jaterní fibrogeneze také tím, že inhibují matrixovou metaloproteinázu-9 a aktivují tkáňový inhibitor matrixových metaloproteináz I. typu (12) .
V genech pro TNF-a a IL-1β byly popsány polymorfismy ovlivňující jejich expresi in vitro. Jednonukleotidové polymorfismy (SNP) v pozici -863 a -238 promotoru TNF-A nezávisle zvyšují transkripci tohoto genu (13,14) . Genový cluster interleukinu 1 na chromosomu 2 obsahuje geny IL-1B a IL1-RN kódující IL-1β a receptorového antagonistu IL- Ira (15) . Haplotyp [-511C; -31T] promotoru IL-1B zvyšuje transkripci genu (16) . Druhý intron IL1-RN obsahuje variabilní počet tandemových repetic (VNTR) délky 86-bp. Alela 2 (IL-1RN*2) zvyšuje koncentraci IL-1β in vitro (17) .
Nosiči alely TNF-A -238A s hepatitidou C mají vyšší riziko rozvoje cirhózy (18) , avšak výsledky studií asociace této alely s ALC jsou kontroverzní (19,20) . Alela IL-1B -31T zvyšuje riziko karcinomu žaludku a u pacientů s hepatitidou C zvyšuje riziko hepatocelulárního karcinomu (21,22) . V souvislosti s ALC dosud nebyla studována. Alela IL-1B -511C zvyšuje riziko ALC (23) v asijské populaci. Asociace alely TNF-A -863A s ALC nebyla studována.
Cílem studie bylo ověřit hypotézu, zda výše uvedené polymorfismy v klíčových genech endotoxinové dráhy patogeneze ALC ovlivní riziko rozvoje ALC.
MATERIÁL A METODIKA
Pacienti a kontroly
Do studie jsme zařadili 100 pacientů kavkazského původu s ALC postupně hospitalizovaných na Klinice hepatogastroenterologie IKEM od března 2004 do prosince 2005. Práh konzumace etanolu byl 40 gramů denně pro ženy a 60 gramů denně pro muže po dobu 10 let. Kontrolní skupinu tvořilo 180 neabstinujících dobrovolníků bez jaterního onemocnění, kteří dříve participovali v epidemiologické studii MONICA (24) . Studie byla schválena etickou komisí a všichni pacienti podepsali informovaný souhlas se zařazením do studie.
Jaterní cirhóza byla diagnostikována na základě klinického vyšetření, které zahrnovalo nález fyzikální, laboratorní, sonografický a nález při gastroskopii. Diagnóza byla stanovena na základě laboratorního průkazu jaterní dysfunkce v kombinaci s klinickými, sonografickými a gastroskopickými známkami portální hypertenze (tj. ascites, ikterus, jaterní encefalopatie, jícnové varixy či portální gastropatie). U 11 pacientů byla diagnóza stanovena teprve na základě výsledku necílené jaterní biopsie, která u všech 11 nemocných prokázala cirhózu s polymorfonukleárním zánětlivým infiltrátem a Malloryho hyalinem. Pacienti s nádorem jater nebo s pozitivitou markerů hepatitidy B nebo C, antinukleárních nebo antimitochondriálních protilátek nebyli do studie zařazeni.
Medián věku pacientů s ALC (N = 100) byl 52,6 (26-72) let, 72 pacienti byli muži. Medián denního příjmu etanolu byl 100 (interkvartilový rozptyl 75-174) gramů. Medián věku kontrol (N = 180) byl 47,9 (24-73) let, 142 kontroly byli muži, medián denní konzumace etanolu činil 21,5 (interkvartilový rozptyl, 14-40) gramů. Mezi skupinami nebyly kromě onemocnění jater klinicky významné rozdíly.
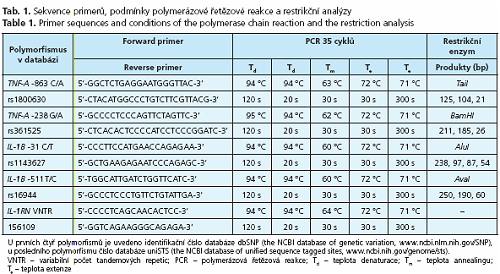
Genotypizace
Genotypizace byla provedena metodou polymerázové řetězové reakce - polymorfismů délky restrikčních fragmentů (PCR-RFLP) (tab. 1). Polymorfismus IL-1RN VNTR byl genotypován metodou PCR-FLR Produkty PCR byly separovány polyakrylamidovou elektroforézou a barveny etidiumbromidem. Restrikční enzymy byly zakoupeny od firmy Fermentas UAB (Vilnius, Litva).
Statistické metody
Velikost souboru byla stanovena výpočtem síly testu při pravděpodobnosti chyby I. druhu 0,05 a chyby II. druhu 0,2, což umožňuje detekci zvýšení relativního rizika (odds ratio, OR) pro ALC více než 1,55 při frekvenci rizikové alely přesahující 0,413 nebo detekci OR > 3 při frekvenci rizikové alely přesahující 0,024. Nezávislá distribuce alel v souboru byla ověřena testem Hardy-Weinbergovy rovnováhy (HWE). Fáze alel v haplotypech byla určena algoritmem maximálně věrohodného odhadu (expectation-maximization, EM). Vazebné dysequilibrium (linkage disequilibrium, LD) mezi polymorfismy na stejném chromosomu (syntenní polymorfismy) bylo charakterizováno koeficienty D' a r2, které nabývají hodnot intervalu <0,1&glt; a určují sílu LD. Hodnota D' = 1 udává kompletní LD při absenci jednoho haplotypu a hodnota r2 = 1 udává absolutní LD při absenci dvou haplotypu ze čtyř možných v případě dialelických SNR Asociační analýza polymorfismů byla provedena x2 testem. Asociace haplotypu s ALC byla analyzována logistickou regresí v programu SPSS, verze 14.0 (SPSS Inc., Chicago, IL, USA). Hladina signifikance pro asociaci byla P < 0,05. Populační atributabilní riziko (PAR) polymorfismů asociovaných s ALC bylo vypočteno dle rovnice:
PAR(%) =((fp - fk) / fp) x 100
kde je PAR populační atributabilní riziko, fp frekvence polymorfismu ve skupině pacientů a fk frekvence polymorfismu ve skupině kontrol.
VÝSLEDKY
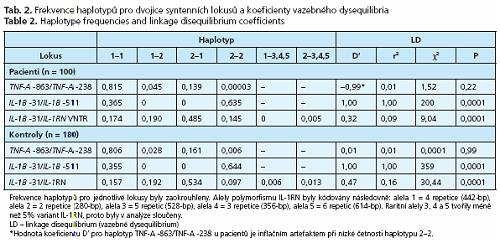
Alely pro jednotlivé lokusy v kontrolní populaci byly v HWE. Lokus TNF-A -863 jako jediný vybočoval z rovnováhy v důsledku nadměrného počtu homozygotů při nízké frekvenci rizikové alely -863A a z asociační analýzy musel být vyřazen. LD mezi lokusy IL-1B -511a IL-1B -31 bylo absolutní (D' = 1, r2 = 1) s výlučnou přítomností haplotypů T-C nebo C-T (tab. 2). Mezi lokusy IL-1B -31 a IL-1RN bylo LD slabé (D' < 0,5). Alely v lokusech TNF-A -863 a TNF-A -238 segregovaly nezávisle.
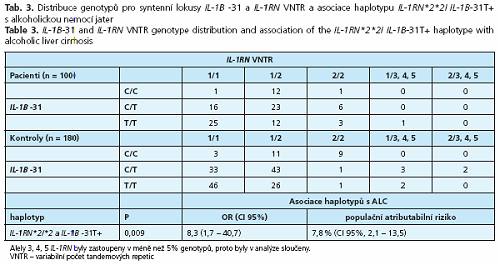
Žádná z alel TNF-A -863A, TNF-A -238A, IL-1B -511C, IL-1B -31T aILl-RN *2 nebyla samostatně asociována s ALC. Haplotyp zahrnující homozygotní stav pro alelu IL-1RN*2 (IL-1RN*2*2) a současně nosičství alespoň jedné alely IL-1B -31T (IL-1B -31T+), zastoupený u 9 % pacientů a u 1 % kontrol, byl asociován s vyšším rizikem ALC (P = 0,009) (tab. 3). Populační atributabilní riziko (PAR) haplotypů IL-1RN *2*2I IL-1B -31T+ určující, do jaké míry determinuje haplotyp ve studované populaci fenotyp, bylo 7,8 %.
DISKUZE
Polymorfismus IL-1B -31C/T se nachází v TATA boxu promotoru. Záměna cytosinu za thymin zvyšuje transkripční aktivitu genu in vitro pětkrát (16) . Lokus IL-1B -31 kosegreguje s lokusem -511 z více než 98% (98,5% El Omar et al. (16) , 100% naše data a databáze HapMap). Jelikož ke kosegregaci dochází ve 100% i v japonské populaci (databáze Hap-Map), lze asociaci mezi rizikovou alelou IL-1B -511 a ALC popsanou ve studii Takamatsu et al. (23) vysvětlit kosegregaci s funkčně relevantní alelou IL-1B -3 IT. V rozporu s výsledkem této studie jsme v našem souboru asociaci alely IL-1B -511C resp. IL-1B -31T s ALC nenalezli. Vysvětlení spočívá v tom, že hodnota PAR pro alelu IL-1 -511C ve studii Takamatsu et al. (23) pro polygenní chorobu vyšla neobvykle vysoká (vyšší než 32%), což je dáno výběrem kontrolní skupiny zdravých dobrovolníků s prokázaným abúzem alkoholu. Při srovnání frekvence rizikové alely IL-1B -511C u nemocných s ALD a v reprezentativním vzorku japonské populace nebyla asociace potvrzena.
Lokus IL-1B -31 je v LD i s alelou IL-1RN*2, u které Santtila et al.(17) prokázal, že zvyšuje produkci IL-1β in vitro, přičemž alela IL-1RN*2 zvyšuje produkci IL-1β více než alela IL-1B -31T Přítomnost LD s IL-1B -31 dokazuje paralelně se studií (25) podíl obou lokusů na sekreci IL-1β. Nízká síla LD (tj. neúplná kosegregace obou lokusů) umožňuje aditivitu vlivu alel na sekreci IL-1β.
Obratem ve studiích genového clusteru IL-1 byla práce El Omar et al. (16) , který zjistil asociaci alel IL-1B -31T a IL-1RN*2 s karcinomem žaludku. Riziko rozvoje onemocnění zvýšila každá alela samostatně, významněji však v haplotypů IL-1RN*2*2I IL-1B -31T+. Haplotypizační analýza našeho souboru logicky navazující na citovanou studii El Omara et al. ukázala, že zmíněný haplotyp se podílí i na dědičné dispozici k rozvoji ALC.
Asociaci alely TNF-A -238A s ALC jsme v naší studii neprokázali. Výsledek je v souladu s negativním nálezem Ladero et al.(20), avšak v rozporu se studiemi Grove et al. (19) a Pastor et al. (26) , které asociaci prokázaly. K pozitivní asociaci popsané v posledně jmenovaných studiích zřejmě významnou měrou přispěl nedostatečný počet pacientů (27) .
Výsledky naší studie ukazují na pravděpodobnou úlohu IL-1β v rozvoji alkoholické cirhózy jater, v jejíž patogenezi hraje významnou úlohu chronická infiltrace polymorfonukleárními lymfocyty vyvolaná působením IL-1β (9) . Trvá-li tato infiltrace léta a je-li doprovázena kontinuálním rozpadem velkého množství granulocytů, přispívá k chronickému postižení jater i expozice jaterního parenchymu lyzosomálním enzymům z rozpadlých granulocytů (10,11) . Tento mechanizmus je patrně odlišný od mechanizmu akutní alkoholické hepatitidy kde hraje klíčovou úlohu apoptóza vyvolaná TNF-ct, na které se IL-1β podstatně nepodílí.
Význam genových polymorfismů v predispozici k ALC je ovlivněn nejen funkcí, ale i frekvencí. Zatímco frekvence rizikových alel IL-1B -31T a IL-1RN*2 činí 0,6 a 0,2, byla často studovaná ale-la TNF-A -238A s frekvencí 0,03 z genofondu téměř eliminována. Přes prokázaný funkční význam polymorfismů TNF-A -238G/A in vitro se v predispozici k ALC více uplatní podstatně častější rizikový haplotyp v genovém clusteru interleukinu 1, zatímco ovlivnění rizika ALC alelou TNF-A -238A nelze v populačních studiích běžného rozsahu dokázat.
ZÁVĚR
Naše výsledky prokazují osminásobné zvýšené rizika rozvoje ALC u homozygotů pro alelu IL-1RN*2, kteří jsou alespoň heterozygoty pro alelu IL-1B -31T Haplotyp IL-1RN*2*2IIL-1B -31T+ představuje 7,8 % dědičné dispozice k rozvoji ALC. Synergický účinek alel IL-1B -3 IT a IL-1RN*2 zvyšuje produkci IL-1β, což potencuje zánětlivou aktivitu a progresi fibrózy v játrech a vysvětluje biologickou podstatu zvýšeného rizika ALC u nosičů tohoto haplotypu.
Literatura
- 1. Corrao G, et al. Trends of liver cirrhosis mortality in Europe, 1970-1989: ageperiodcohort analysis and changing alcohol consumption. Int J Epidemiol 1997; 26: 100-109.
- 2. Stickel F, Osterreicher CH. The role of genetic polymorphisms in alcoholic liver disease. Alcohol Alcohol 2006; 41: 209-224.
- 3. Hrubec Z, Omenn GS. Evidence of genetic predisposition to alcoholic cirrhosis and psychosis: twin concordances for alcoholism and its biological end points by zygosity among male veterans. Alcohol Clin Exp Res 1981; 5: 207-215.
- 4. Reed T, et al. Genetic predisposition to organspecific endpoints of alcoholism. Alcohol Clin Exp Res 1996; 20: 1528-1533.
- 5. McClain CJ, et al. Cytokines in alcoholic liver disease. Semin Liver Dis 1999; 19: 205-219.
- 6. Natori S, et al. Hepatocyte apoptosis is a pathologic feature of human alcoholic hepatitis. J Hepatol 2001; 34: 248-253.
- 7. Felver ME, et al. Plasma tumor necrosis factor alpha predicts decreased long-term survival in severe alcoholic hepatitis. Alcohol Clin Exp Res 1990; 14: 255-259.
- 8. Kanzler S, Galle PR. Apoptosis and the liver. Semin Cancer Biol 2000; 10: 173-184.
- 9. Bautista AP. Neutrophilic infiltration in alcoholic hepatitis. Alcohol 2002; 27: 17-21.
- 10. Sauer A, et al. Endotoxin-induciblegranulocyte-mediated hepatocytotoxicity requires adhesion and serine protease release. J Leukoc Biol 1996; 60: 633-643.
- 11. Colombat M, et al. Portal lymphocytic infiltrate in alcoholic liver disease. Hum Pathol 2002; 33: 1170-1174.
- 12. Roderfeld M, et al. Cytokine blockade inhibits hepatic tissue inhibitor of metalloproteinase-1 expression and up-regulates matrix metalloproteinase-9 in toxic liver injury. Liver Int 2006; 26: 579-586.
- 13. Udalova IA, et al. Functional consequences of a polymorphism affecting NF-kappaB p50-p50 binding to the TNF promoter region. Mol Cell Biol 2000; 20: 9113-9119.
- 14. Bayley JP, et al. Functional analysis of linker-scan mutants spanning the -376, -308, -244, and -238 polymorphic sites of the TNF-alpha promoter. Cytokine 2001; 14: 316-323.
- 15. Dinarello CA. Biologic basis for interleukin-1 in disease. Blood 1996; 87: 2095-2147.
- 16. El Omar EM, et al. Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature 2000; 404: 398-402.
- 17. Santtila S, Savinainen K, Hurme M. Presence of the IL-1RA allele 2 (IL1RN*2) is associated with enhanced IL-lbeta production in vitro. Scand J Immunol 1998; 47: 195-198.
- 18. Yee LJ, et al. Tumor necrosis factor gene polymorphisms in patients with cirrhosis from chronic hepatitis C virus infection. Genes Immun 2000; 1: 386-390.
- 19. Grove J, et al. Association of a tumor necrosis factor promoter polymorphism with susceptibility to alcoholic steatohepatitis. Hepatology 1997; 26: 143-146.
- 20. Ladero JM, et al. Single nucleotide polymorphisms and micro-satellite alleles of tumor necrosis factor alpha and interleukin-10 genes and the risk of advanced chronic alcoholic liver disease. Liver 2002; 22: 245-251.
- 21. El Omar EM, et al. Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology 2003; 124: 1193-1201.
- 22. Wang Y, et al. Interleukin-lbeta gene polymorphisms associated with hepatocellular carcinoma in hepatitis C virus infection. Hepatology 2003; 37: 65-71.
- 23. Takamatsu M, et al. Genetic polymorphisms of interleukin-lbeta in association with the development of alcoholic liver disease in Japanese patients. Am J Gastroenterol 2000; 95: 1305-1311.
- 24. Multinational Monitoring of Trends and Determinants in Cardiovascular Diseases: „MONICA Project." Manual of operations, WHO/MNC 82.2.
- 25. Hirsch E, et al. Functions of interleukin 1 receptor antagonist in gene knockout and overproducing mice. Proc Natl Acad Sci USA 1996; 93: 11008-11013.
- 26. Pastor IJ, et al. -238 G>A polymorphism of tumor necrosis factor alpha gene (TNFA) is associated with alcoholic liver cirrhosis in alcoholic Spanish men. Alcohol Clin Exp Res 2005; 29: 1928-1931.
- 27. Wilhelmsen KC. Does tumor necrosis factor play a role in alcoholic steatohepatitis? The potential pitfalls of a case-controlled allelic association analysis. Hepatology 1997; 26: 232-233.
Pro přístup k článku se, prosím, registrujte.
Výhody pro předplatitele
Výhody pro přihlášené